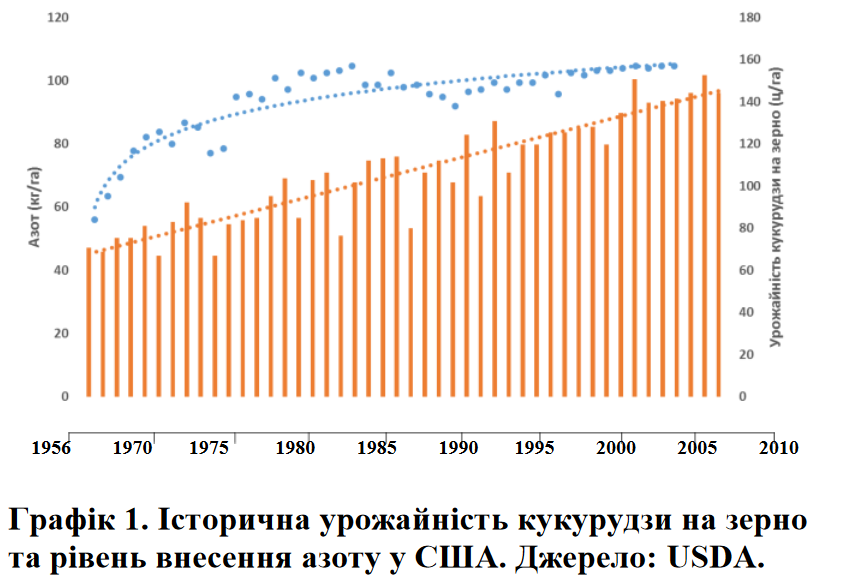
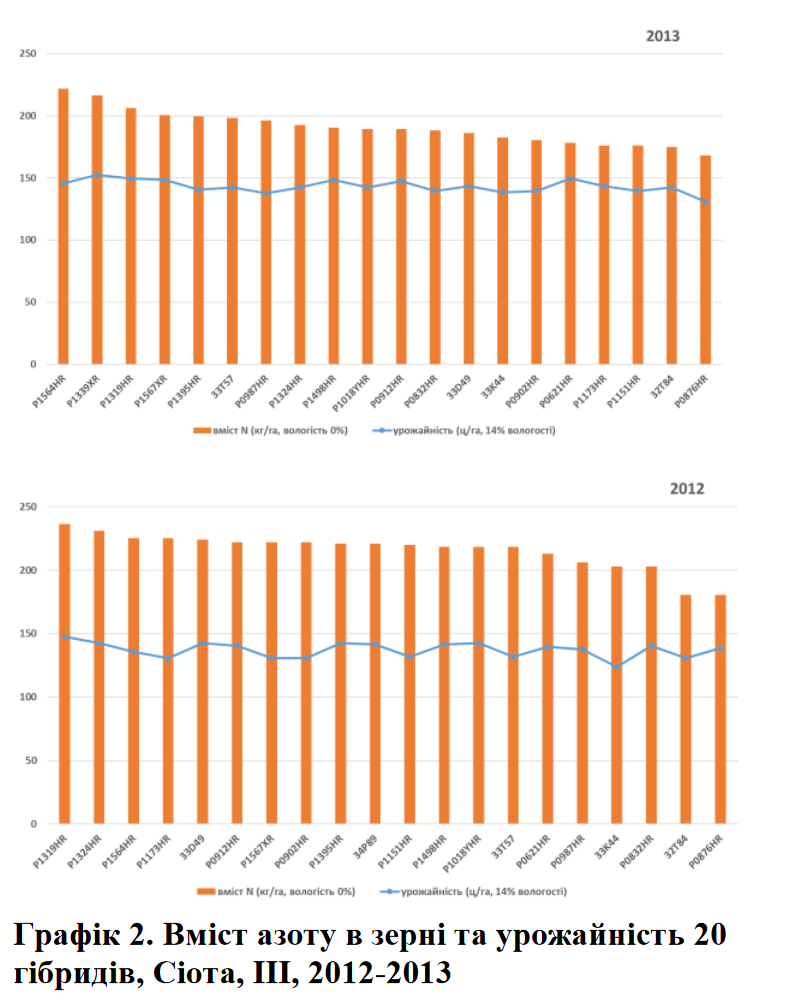
Важливість азотного живлення до цвітіння важко переоцінити, оскільки азот підтримує формування початків, закладку кількості зерен та їх розмір. Через це аграрії зазвичай намагалися забезпечити доступність азоту саме на початковому етапі вегетації, вважаючи внесення азоту після цвітіння не дуже важливим. Історичні дослідження по часу внесення азоту також підкреслювали важливість його раннього внесення. Більше того, переважна більшість досліджень стверджувала, що при наливі зерна майже вся потреба в азоті покривалася за рахунок його перерозподілу із листя та стебел до зернівок, зводячи до мінімуму вбирання азоту з грунту після цвітіння.
Однак всебічні дослідження останніх 5 років довели, що азот, необхідний для розвитку зерна, має 2 джерела походження:1) перерозподіл із листя та стебел; 2) забір азоту із грунту. До такого висновку прийшли як науковці Corteva, так і представники дослідницьких інститутів.
Дослідження Cortevaу Макомбі, ІІІ у 2012 році порівнювали «звичайний» рівень азоту – 224 кгN/га із низьким рівнем –56 кгN/га. Отримані врожаї склали 157 ц/га при нормальному забезпеченні та 63 ц/га при низькому. При нормальному забезпеченні азотом 145, 180 та 336 кгN/га вбиралося на момент V12, R1 та R6 відповідно (графік 4). Із 224 кгN/га, що містилося в зерні на момент фізіологічної стиглості, 68 кг походили із вегетативних органів рослини (листя, стебла, тощо) і 156 кг було отримано із грунту після цвітіння (графік 4).
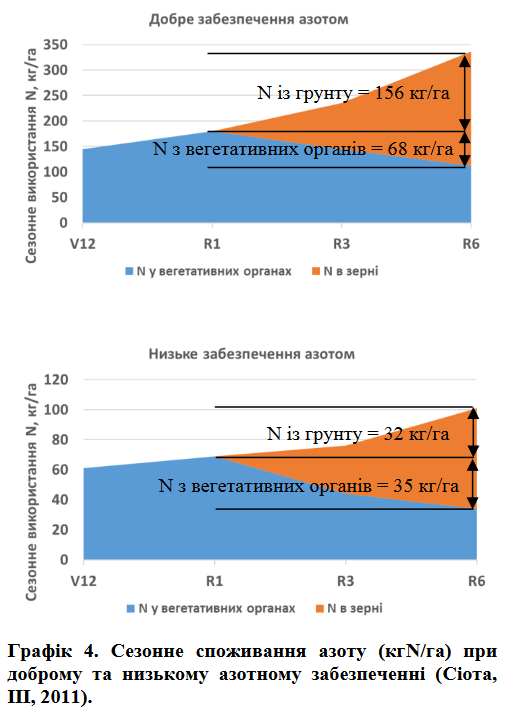
При низькому азотному живленні (урожайність на рівні 63 ц/га) споживання азоту на стадії R1 обмежувалося 69 кг/га і тільки 32 кг/га вбиралося із грунту після цвітіння (графік 4). Низьке забезпечення азотом імітує умови, коли доступний N обмежується вилуговуванням, денітрифікацієюта недостатнім його внесенням.
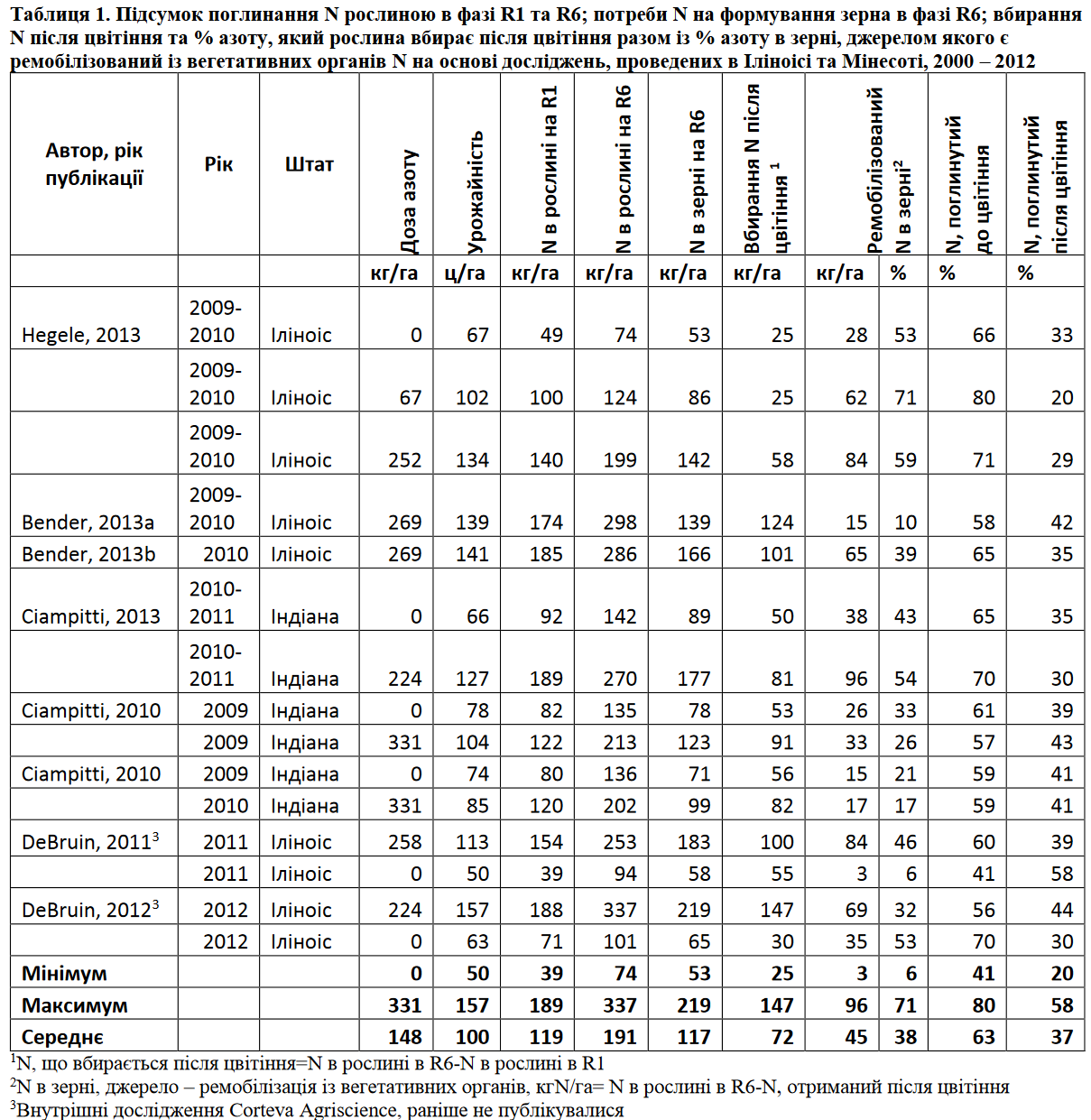
В умовах низького азотного живлення, коли вбирання N після цвітіння не може в повному обсязі забезпечити формування зерна, решта азоту ремобілізується із вегетативних органів (стебла, листя, обгортки, стрижень). Листя –основне джерело ремобілізованого азоту. Сучасні дослідження, проведені в Іліноісі та Індіані, показали, що в середньому доля ремобілізованого із вегетативних органів азоту складає 38%, а максимальне значення – 54% –має місце у сприятливих грунтово-кліматичних умовах із високим азотним живленням (таблиця 1). Незалежні дослідження ДеБруіна та інших (2012) зафіксували 63% як максимально можливий рівень ремобілізації азоту в зерно. Подальші дослідженнянауковців Cortevaдовели, що кількість ремобілізованогоіз стебел азоту складає близько 20%, а перерозподіл азоту із обгорток та стрижня в зерно є дуже незначним.
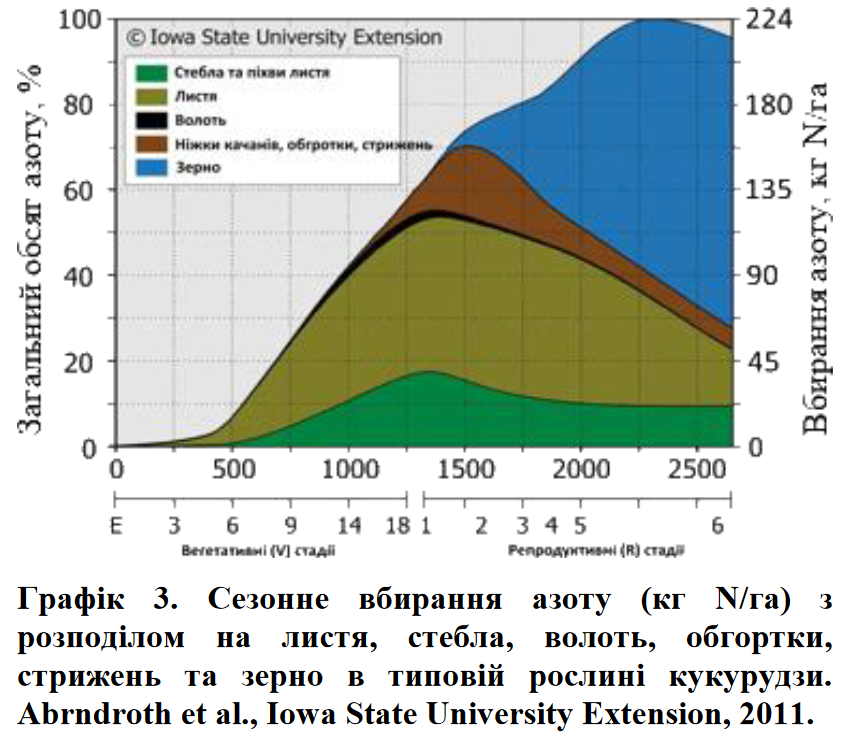
Декілька університетських досліджень оцінювали час вбирання азоту. В дослідженніДержавного Університету Айови приблизно 60% (135кгN/га) від загальної потреби в азоті (графік 3) було ввібрано і накопичено в листі, стеблах та початках на момент R1 (вихід шовку) в умовах сприятливих грунтово-кліматичних умов –рівень урожайності 140 ц/га (Abendroth et al., 2011). На момент фізіологічної стиглості (R6 –чорна точка) –загальний обсяг ввібраного азоту (вегетативні органи+зерно) досягав 213 кгN/га. Подальші розрахунки показують, що 80 кгN/га все ще повинні бути поглинуті після цвітіння для підтримки формування зерна.
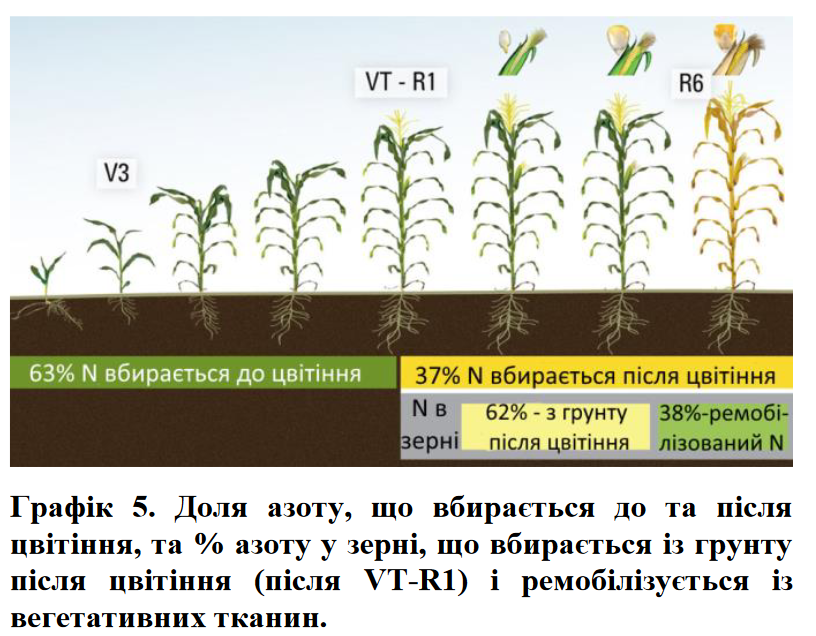
Результати інших досліджень щодо пропорції азоту, що споживається до і після цвітіння, наведені на графіку 5 та в таблиці 1.Вони доводять, що кількість азоту, використаного до цвітіння, практично ідентична в умовах доброго і низького азотного живлення (63% та 62% відповідно). Незважаючи на те, що це – значна частка сезонної потреби в азоті, потреба в N після цвітіння коливається від 36 до 150 кгN/га.
Приклади важливості вбирання азоту після цвітіння наведені у таблиці 1. У 2 різних середовищах в Іліноісі і Індіані з рівнями урожайності більше 140 ц/га рівень поглинання азоту до R1 досягнув 188 кгN/га. Оскільки максимально можливе значення ремобілізованого азоту складає 54% від обсягу, що поглинувся до цвітіння, тільки 100 кгN/га буде доступне для наливу зерна. Припускаючи вміст білку в зерні на рівні 8% ця кількість азоту буде достатньою для формування урожаю тільки на рівні 80 ц/га. Таким чином, рівень урожайності кукурудзи буде дуже обмежений в разі використання для наливу зерна тільки ремобілізованого азоту, особливо це стосується полів, де наявність азоту зменшується за рахунок грунтово-кліматичних умов та через постійне недовнесення. Із таблиці 1 чітко видно, що сучасні гібриди в значній мірі залежать від вбирання азоту із грунту після цвітіння для успішного наливу зерна.
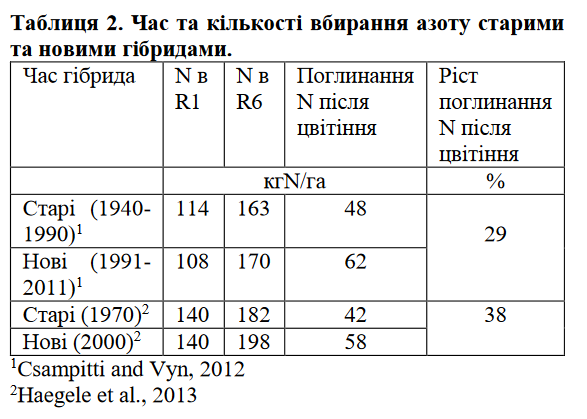
Одна з причин непорозумінь щодо поглинання азоту після цвітінні і його впливу на налив зерна є думка, що сучасні, високоврожайні гібриди мають відмінний тренд вбирання N. Сучасними дослідженнями доведено, що гібриди сучасної селекції поглинають більші кількості N після цвітіння в порівнянні із старішими гібридами. Дослідження Ciampitti та Vyn (2012) узагальнили 100 наукових праць по старим (1940-1990) і новим (1991- 2011) гібридам (таблиця 2). Науковці прийшли до висновку, що в середньому сучасні гібриди вбирали на 3 кг N більше протягом вегетаційного періоду. Ще більш важливим є час поглинання N рослинами: нові гібриди поглинають на 29% більше азоту після цвітіння в порівнянні із старими гібридами (таблиця 1).
Haegele (2013) поглибив дослідження щодо часу вбирання азоту рослинами кукурудзи. Він порівняв гібриди із 1970-х років із гібридами, виведеними після 2000 року (таблиця 2). Згідно його досліджень, сучасні гібриди поглинали на 3.6 кг N більше протягом вегетаційного сезону і акумулювали на 40% більше азоту після цвітіння в порівнянні із старими гібридами (таблиця 2). Оцінка гібридів Pioneer, які продавалися у 1934 – 2013 роках дала аналогічні результати: сучасні гібриди поглинають більше азоту протягом вегетаційного періоду, а по часу, в порівнянні із старими гібридами, більша частина поглинається після цвітіння. Враховуючи наведені напрацювання, варто зробити переоцінку рекомендацій щодо часу внесення азоту та підтримки достатнього рівня азотного живлення протягом репродуктивних фаз розвитку кукурудзи.