Corn Rootworm Management Using RNAi
Something went wrong. Please try again later...
Corn rootworm has long been one of the most damaging insect pests of corn in North America. There are four rootworm species that affect corn: western corn rootworm (Diabrotica virgifera virgifera), northern corn rootworm (D. barberi), Mexican corn rootworm (D. virgifera zeae), and southern corn rootworm (D. undecimpunctata howardi). Of these four species, the western and northern corn rootworm are the most economically important and the most challenging to control.
Western and northern corn rootworms have a history of adapting to and overcoming control practices, which has increased the complexity and difficulty of successfully managing these pests.
Crop rotation: Historically, crop rotation was an effective and widely used management strategy, and it is still a very important part of an integrated management strategy; however, populations of both western and northern corn rootworm have developed the ability to survive in two-year corn-soybean rotations.
Insecticides: Resistance to several classes of insecticides has been documented in populations of western corn rootworm, both to soil applications for larva control and foliar applications for adult control.

Bt Traits: Bt corn hybrids engineered to express genes from the common soil bacterium Bacillus thuringiensis have been an important management tool for corn rootworm for the past 20 years. However, field-evolved resistance in western corn rootworm has now been documented for all four Bt traits for corn rootworm protection currently on the market (Table 1).
Cross-resistance between the Cry3 proteins (Cry3Bb1, mCry3A, and eCry3.1Ab) has been demonstrated, which means that the four current Bt traits only provide two effective modes of action against western corn rootworm (Jakka et al., 2016).
Table 1. Bt technologies currently on the market for protection against corn rootworm and year that field-evolved resistance in western corn rootworm was first documented.
Bt Protein |
Original |
First Case of Field-Evolved Resistance |
Cry3Bb1 |
YieldGard® Rootworm |
2011a |
Cry34/35Ab1 |
Herculex® RW |
2013b |
mCry3A |
Agrisure® RW |
2014c |
eCry3.1Ab |
Agrisure Duracade® |
2016d |
aGassmann et al., 2011;
bGassmann et al., 2016;
cGassmann et al., 2014;
dZukoff et al., 2016
The destructiveness of western corn rootworm in corn and its continuing ability to overcome management tactics has created an urgent need for additional management tools. Ribonucleic acid interference (RNAi) technology has been commercialized to provide an additional unique mode of action for protection against corn rootworm. The first transgenic corn product with an RNAi-based plant incorporated protectant for corn rootworm management was registered by the U.S. EPA in 2017. This product includes two Bt traits (Cry3Bb1 and Cry 34/35Ab1) plus RNAi to provide a total of three effective modes of action against corn rootworm. This combination of three modes of action is available in Corteva Agriscience seed brands as Vorceed™ Enlist® corn.
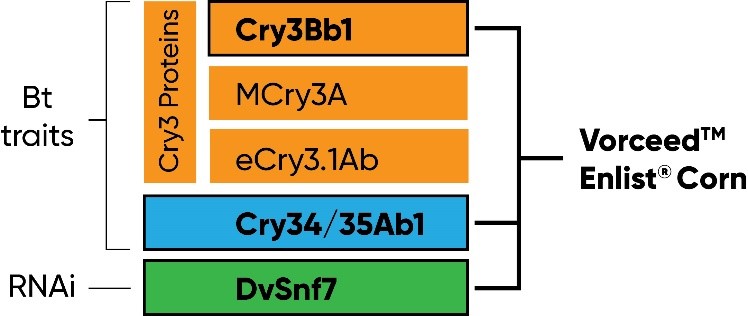
Figure 1. Vorceed Enlist corn contains three modes of action for protection against corn rootworm: two Bt traits (Cry3Bb1 and Cry34/35Ab1) and RNAi (DvSnf7).
RNA interference (RNAi) is a mechanism in cells that regulates gene expression by reducing or "silencing" the activity of specific genes. It does this by using small RNA molecules called siRNAs (short interfering RNAs) to target and degrade the messenger RNA (mRNA) molecules that code for the gene of interest. The RNAi mechanism is found in many organisms, from plants and animals to fungi and bacteria.
RNAi is thought to have evolved as a defense mechanism against invasive genetic elements such as RNA viruses and transposable elements (mobile DNA sequences that can replicate and insert themselves into different locations within the host genome, also known as “jumping genes”). The RNAi gene-silencing mechanism was first discovered in the roundworm Caenorhabditis elegans in 1998 (Fire et al., 1998); a discovery that was awarded a Nobel Prize in 2006.
The potential utility of RNAi as a mechanism to protect corn against corn rootworm was first demonstrated in 2007 (Baum et al., 2007).
The process of RNA interference starts with the production of double-stranded RNA (dsRNA) molecules that match the sequence of the target gene (Figure 2). These dsRNA mole-cules are recognized by an enzyme that cuts them into short pieces (siRNAs). The siRNAs are then loaded onto a protein complex called the RNA-induced silencing complex (RISC).
Once the RISC is loaded with the siRNAs, it can search the cell for mRNAs that have a complementary sequence to the siRNAs. When the RISC complex finds an mRNA molecule that matches the siRNA, it cleaves the mRNA, leading to its degradation. This prevents the mRNA from being translated into a protein, effectively silencing the gene.
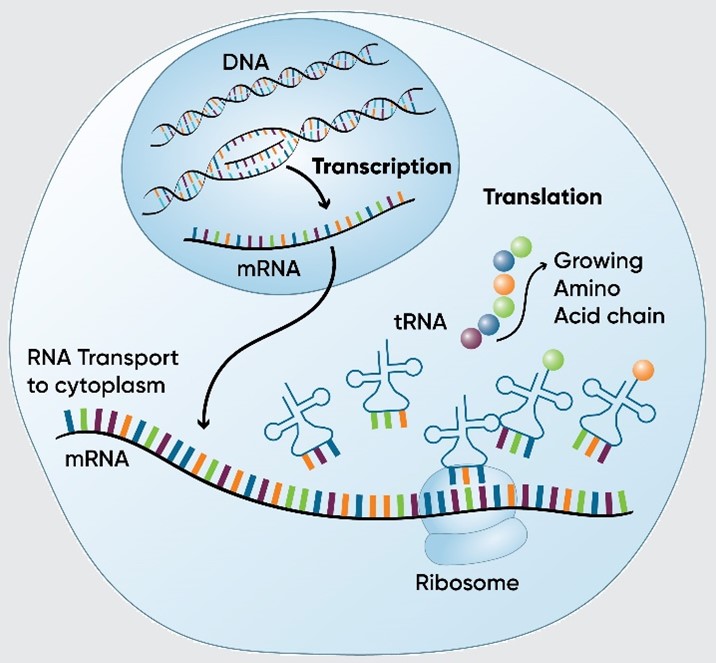
There are two primary processes involved in producing proteins from genetic information coded in DNA: transcription and translation.
Transcription is the process by which genetic information stored in DNA is used to produce a complementary messenger RNA (mRNA) molecule. It occurs in the nucleus of eukaryotic cells.
Translation is the process by which the genetic information carried by mRNA is decoded to produce a specific sequence of amino acids, which form a protein. It occurs in the cytoplasm of all cells.
RNAi is a form of post-transcriptional gene silencing (PTGS), which refers to regulation of gene expression that occurs after transcription has taken place and involves the degradation or inhibition of mRNA.
The RNAi technology in Vorceed™ Enlist® corn protects against corn rootworm by silencing the Snf7 gene, a gene in the corn rootworm genome that codes for a protein necessary for its survival. Snf7 is one of several RNAi target genes that has been evaluated for control of corn rootworm. An ideal RNAi target gene is one that is involved in a critical physiological process in the insect pest – so that silencing it will lead to insect mortality – and one for which the dsRNA expressed by the crop does not affect non-target organisms.
Gene silencing via the RNAi pathway involved three steps: dsRNA uptake, gene silencing, and systemic spread.
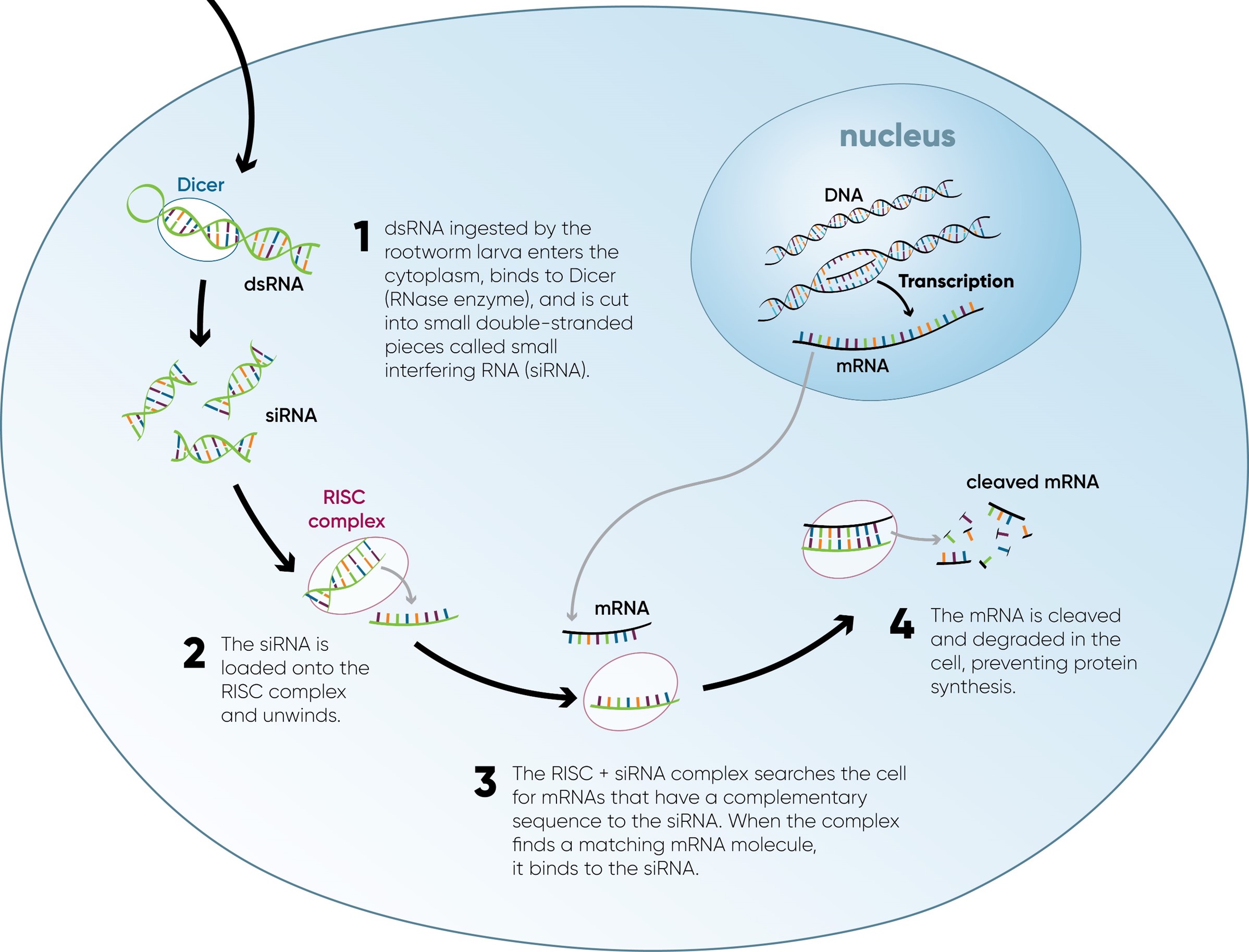
Figure 2. Diagram showing the key steps in the RNAi pathway in a corn rootworm cell. dsRNA produced by the corn plant and ingested by the corn rootworm initiates the RNAi process in the rootworm’s cells, blocking production of a key protein essential for the rootworm’s survival.
Uptake: A gene is inserted into the corn genome that codes for a corn rootworm Snf7 homolog (DvSnf7). When this DNA is transcribed in the corn plant, the resulting RNA folds onto itself forming dsRNA. The DvSnf7 dsRNA is ingested by corn rootworm larvae when they feed on the corn roots. Once ingested, the dsRNA initiates the RNAi process in the rootworm’s cells.
Gene Silencing: The dsRNA is cleaved into siRNAs, which are then loaded onto the RISC. The RISC then targets Snf7 mRNA molecules, binds to them, and cuts them up; effectively silencing the gene by preventing the mRNA from being translated into a protein. Since the corn rootworm needs this protein to survive, deprivation eventually leads to larval death.
Systemic Spread: Following ingestion by a corn rootworm larva, the dsRNA moves beyond the gut of the insect and spreads systemically throughout the body. This systemic spread is crucial in achieving sufficient gene silencing to kill the larva.
The RNAi technology in Vorceed™ Enlist® corn is active against all four corn rootworm species that impact corn in North America: western, northern, southern, and Mexican corn rootworm. As with Bt proteins, RNAi-based modes of action require corn rootworm larvae to feed on roots and ingest root tissue to be exposed to the insecticidal dsRNA.
RNAi can result in insect mortality comparable to that of a Bt protein; however, the RNAi-based mode of action generally is slower to kill corn rootworm larvae than Bt proteins due to the multistep process involved in RNAi uptake, gene suppression, and systemic spread. If it were used alone and not stacked with Bt traits, RNAi would significantly reduce the emerging adult rootworm population but would not provide adequate root protection due to the longer time to mortality. Because of this – as well as the need to promote trait durability – DvSnf7 is not marketed as a single trait product and is only available in combination with at least one Bt trait.
Field trials showed that the addition of DvSnf7 dsRNA to corn hybrids with Cry3Bb1 and Cry34/35Ab1 Bt traits significantly reduced root damage in fields with high western corn rootworm densities compared to corn with only the Bt traits (Head et al., 2017). The addition of DvSnf7 dsRNA to Bt hybrids was also effective in reducing western corn rootworm adult emergence. DvSnf7 dsRNA is expressed throughout the corn plant – including plant tissues commonly fed upon by corn rootworm adults – however, concentrations are not high enough to cause mortality in adults.
Baum, J.A., T. Bogaert, W. Clinton, G.R. Heck, P. Feldmann, O. Ilagan, S. Johnson, G. Plaetinck, T. Munyikwa, M. Pleau, et al. 2007. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 25:1322-1326.
Darlington, M., J.D. Reinders, A. Sethi, A.L. Lu, P. Ramaseshadri, J.R. Fischer, C.J. Boeckman, J.S. Petrick, J.M. Roper, K.E. Narva, et al. 2022. RNAi for Western Corn Rootworm Management: Lessons Learned, Challenges, and Future Directions. Insects 13:57.
Fire, A., S. Xu, M.K. Montgomery, S.A. Kostas, S.E. Driver, and C.C. Mello. 1998. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 391, 806–811.
Gassmann, A.J., J.L. Petzold-Maxwell, R.S. Keweshan, and M.W. Dunbar. 2011. Field-evolved resistance to Bt maize by western corn rootworm. PLOS ONE 6(7): e22629.
Gassmann, A.J., J.L. Petzold-Maxwell, E.H. Clifton, M.W. Dunbar, A.M. Hoffmann, D.A. Ingber, and R.S. Keweshan. 2014. Field-evolved resistance by western corn rootworm to multiple Bacillus thuringiensis toxins in transgenic maize. Proc. Nat. Acad. Sci. 111 (14) 5141-5146.
Gassmann A.J., R.B. Shrestha, S.R.K. Jakka, M.W. Dunbar, E.H. Clifton, A.R. Paolino, D.A. Ingber, B.W. French, K.E. Masloski, J.W. Dounda, and C.R. St. Clair. 2016. Evidence of resistance to Cry34/35Ab1 corn by western corn rootworm (Coleoptera: Chrysomelidae): root injury in the field and larval survival in plant-based bioassays. J Econ Entomol 109: 1872– 1880 (2016).
Head, G.P., M.W. Carroll, S.P. Evans, D.M. Rule, A.R. Willse, T.L. Clark, N.P. Storer, R.D. Flannagan, L.W. Samuel, and L..J. Meinke. 2017. Evaluation of SmartStax and SmartStaxPRO maize against western corn rootworm and northern corn rootworm: Efficacy and resistance management. Pest Manag. Sci. 73:1883-1899.
Jakka, S.R.K., R.B. Shrestha, and A.J. Gassmann. 2016. Broad-spectrum resistance to Bacillus thuringiensis toxins by western corn rootworm (Diabrotica virgifera virgifera) Sci Rep 6, 27860.
Zukoff, S.N., K.R. Ostlie, B. Potter, L.N. Meihls, A.L. Zukoff, L. French. M.R. Ellersieck, B.W. French, and B.E. Hibbard. 2016. Multiple assays indicate varying levels of cross resistance in Cry3Bb1-selected field populations of the western corn rootworm to mCry3A, eCry3.1Ab, and Cry34/35Ab1. J Econ Entomol 109: 1387-1398.
Liberty®, LibertyLink® and the Water Droplet Design are registered trademarks of BASF. Agrisure® is a registered trademark of, and used under license from, a Syngenta Group Company.
The foregoing is provided for informational use only. Please contact your Pioneer sales professional for information and suggestions specific to your operation. Product performance is variable and depends on many factors such as moisture and heat stress, soil type, management practices and environmental stress as well as disease and pest pressures. Individual results may vary. CI230503
May 2023